Gli esseri umani sono animali sociali capaci di elaborare e capire le azioni e le intenzioni degli altri, questa capacità è di primaria importanza per poter agire ed interagire correttamente e in maniera adattiva con il mondo, qual’è il ruolo dei neuroni specchio?
Beatrice Agostini – OPEN SCHOOL, Psicoterapia Cognitiva e Ricerca Bolzano
Immaginiamo di camminare per strada e di vedere un signore che si avvicina muovendo una mano. Essere in grado di capire se quel movimento è un movimento di saluto o se il signore in questione ci sta per lanciare addosso qualcosa è di primaria importanza per pianificare correttamente il nostro comportamento (ricambiare il saluto nel primo caso, scappare o attaccare nel secondo).
Neuroni specchio: la scoperta
Negli anni ’90 i ricercatori iniziarono ad interrogarsi su come il nostro cervello riconosce le azioni altrui. Nel 1992, di Pellegrino e colleghi studiarono un gruppo di neuroni localizzati nella parte rostrale della corteccia premotoria ventrale del cervello della scimmia (area F5) ed osservarono come questi neuroni si attivavano non solo quando la scimmia faceva un movimento, ma anche quando osservava lo stesso movimento eseguito dallo sperimentatore (Rizzolatti et al., 1996). Questi neuroni vennero chiamati “neuroni specchio”, in inglese “mirror neurons”, proprio per enfatizzare questa loro capacità di rispecchiare una specifica azione motoria nel cervello dell’osservatore.
Studi più approfonditi hanno dimostrato che l’osservazione di azioni altrui determina anche negli esseri umani, e non solo nelle scimmie, l’attivazione delle regioni precentrali (Gallese et al., 2004; Rizzolatti e Craighero, 2004; Keyser e Gazzola, 2009). Questo risultato è stato immediatamente interpretato come parte cruciale del processo di riconoscimento delle azioni: un osservatore comprende le azioni delle altre persone perché le rappresenta nel proprio cervello, proprio come se lui stesso stesse eseguendo quell’azione. Rizzolatti e colleghi (2001) non si fermano qui, ma ipotizzano anche l’esistenza di un network (mirror neuron system), che comprende aree parietali, frontali inferiori e premotorie, che è deputato al riconoscimente delle azioni non solo quando le osserviamo ma anche quando leggiamo un verbo o ascoltiamo una parola associata ad una azione. Ad esempio, un concetto come saltare (indipendentemente se osserviamo una persona che salta, se la immaginiamo o se leggendo un libro incontriamo questa parola) verrebbe compreso grazie alla riattivazione dello stesso programma motorio che si attiverebbe se stessimo effettivamente facendo un salto.
Neuroni specchio: come funzionano?
Iniziò così, verso la metà degli anni ’90, a prendere piede l’idea che le rappresentazioni concettuali riferite alle azioni (ovverso le rappresentazioni semantiche, il significato) siano rappresentate all’interno del nostro sistema sensorimotorio (embodied cognition hypothesis o teoria della cognizione incarnata). In particolare, il concetto di simulazione come il “processo attraverso il quale i concetti rievocano gli stati percettivi e motori presenti quando percepiamo e agiamo nel mondo” (Chatterjee, 2010 –p.80) divenne il focus delle ricerche nell’ambito dell’osservazione e del riconoscimento di azioni (review: Martin, 2007; Mahon e Caramazza, 2008; Kiefer e Pulvermüller, 2012). Lo stesso concetto è stato utilizzato per speculare su altri domini cognitivi come ad esempio l’empatia e il riconoscimento delle emozioni (Spaulding, 2012), la teoria della mente (Gallese e Goldman, 1998, Schulte-Ruchter et al., 2007), e sulla natura di diversi disturbi come l’autismo (Dapretto et al., 2005; Oberman et al., 2005; Hadjikhani et al., 2006).
Ad oggi, non tutti i ricercatori condividono questa interpretazione. Il dibattito su quale sia il ruolo di questo processo di simulazione mentale nel riconoscimento delle azioni è ancora aperto. In particolare ci si domanda: è davvero necessario simulare un’azione nel nostro sistema motorio per comprenderla? Ovvero, l’informazione motoria è fondamentale per comprendere un concetto? In alternativa: è possibile comprendere il significato di una azione solo utilizzando una rappresentazione simbolica, senza il contributo del circuito motorio necessario per metterla in atto? E se si, dove si trova, nel cervello, questa rappresentazione simbolica? Queste sono le domande chiave che caratterizzano il dibattito tra teoria motoria e teoria cognitiva del riconoscimento delle azioni (in qualche misura sovrappoinibili alle embodied/disembodied cognition hypothesis).
Neuroni specchio e teoria motoria del riconoscimento delle azioni
La teoria motoria del riconoscimento delle azioni concorda con la teoria dei neuroni specchio e quindi con la teoria della embodied cognition hypothesis, la quale afferma che la cognizione dipenda anche da caratteristiche di tipo corporeo (nel nostro caso l’informazione contenuta nel sistema motorio). Questa teoria sostiene che l’abilità di capire o riconoscere il significato di un’azione è situata nel nostro sistema motorio. Come si diceva prima, è possibile riconoscere un’azione solo se vi è una simulazione dell’azione osservata nel sistema motorio dell’osservatore.
La maggior parte degli studi comportamentali citati a favore della teoria motoria cercano di dimostrare come le rappresentazioni motorie e le rappresentazioni concettuali interagiscono tra loro, e, soprattutto, come le prime sono in grado di influenzare le seconde. In un esperimento per dimostrare che le parole automaticamente attivano la rappresentazione motoria, Glover e colleghi (2004) mostrarono ai partecipanti il nome di un oggetto grande o piccolo (ad esempio mela o uva). Il compito dei partecipanti era quello di leggere il nome dell’oggetto e subito dopo raggiungere ed afferrare un oggetto target presente sul tavolo (grasping movement). I risultati mostrarono che l’apertura della mano durante il movimento di grasping era influenzato dalla parola che veniva letta in precedenza: se veniva letto il nome di un oggetto grande, i partecipanti aprivano la mano di più rispetto a quando leggevano il nome di un oggetto piccolo. Questo indipendentemente dalla dimensione dell’oggetto target che dovevano afferrare. Questo esperimento, insieme a molti altri (Glenberg e Kaschak, 2002; Brass et al., 2001, Craighero et al., 2002; Tucker e Ellis, 2004; Bub et al., 2008) vennero utilizzati come prova del fatto che il nostro sistema motorio si attiva automaticamente quando leggiamo determinate parole.
Studi più recenti hanno utilizzato la stimolazione magnetica transcranica (TMS) per investigare in maniera più diretta il ruolo del sistema motorio nel riconoscimento delle azioni. Quando si utilizza la TMS si va ad interferire con la normale attività di un’area cerebrale e questo genera un cambiamento nel comportamento del partecipante (Rossini et al., 2015). Solitamente, a seconda della procedura seguita, si può osservare un aumento o una diminuzione dei tempi di reazione del partecipante oppure si possono osservare delle variazioni in alcuni parametri elettrofisiologici, come, ad esempio, nei potenziali motori evocati (MEP). Ad esempio, se applichiamo un elettrodo su un muscolo della mano e poi applichiamo un impulso TMS sull’area motoria (M1) che controlla la mano, osserveremo una contrazione del muscolo della mano. L’intensità della contrazione cambia a seconda dell’eccitabilità corticale: più l’attività dell’area motoria è intensa, maggiore sarà la contrazione. MEP è dunque una misura non invasiva dell’eccitabilità del sistema cortico-spinale e quindi una misura della sensibilità di M1. Utilizzando questo metodo è stato dimostrato che osservare il movimento di una mano provoca nei soggetti un aumento dei MEP nel muscolo della loro mano (Fadiga e Rizzolatti, 1995; Strafella e Paus, 2002; Maeda et al., 2002). Questo significa che l’area motoria dell’osservatore era “attiva” mentre osservava il movimento e che quindi osservare azioni determina una modulazione del sistema motorio.
Altri studi hanno dimostrato che M1 è sensibile non solo all’osservazione ed esecuzione di azioni, ma anche quando abbiamo a che fare con un linguaggio associato ad un’azione (ad es. “spegni la luce”). In un esperimento del 2005, Buccino e colleghi applicarono la TMS sull’area motoria della mano o del piede del partecipante, mentre questo ascoltava delle frasi che si riferivano ad azioni manuali o che coinvolgevano l’uso dei piedi. Durante la stimolazione venivano registrati i MEP. I risultati mostrarono che l’intensità dei MEP del muscolo della mano era diversa quando i partecipanti ascoltavano frasi contenti azioni manuali rispetto a quando ascoltavano frasi che coinvolgevano l’uso dei piedi. L’osservazione che l’area motoria si attiva in maniera specifica quando comprendiamo un’azione, venne interpretata come evidenza del suo coinvolgimento nel riconoscimento semantico delle azioni e venne utilizzata in supporto delle teorie motorie.
Non da ultimo, studi di risonanza magnetica funzionale (fMRI) hanno mostrato che stimoli che fanno riferimento ad azioni motorie (come ad esempio osservare un movimento) portano all’attivazione, tra le altre aree, del giro precentrale (per una review: Martin, 2007; Puvermuller e Fadiga, 2010).
E’ tuttavia importante notare come gli studi sopracitati mostrino una certa variabilità nei risultati. Ad esempio, negli studi di TMS a fronte dello stesso stimolo alcuni studi trovano un aumento dell’intensità dei MEP, altri una diminuzione. In entrambi i casi il risultato è significativo ma la direzione dell’effetto è opposta ed è dunque difficile compararlo o interpretarlo (review: Papeo et al. 2013). Non solo, gli studi di fMRI citati a favore della teoria motoria mostrano sì che il giro precentrale (l’area premotoria) risponde quando elaboriamo il significato delle azioni, ma non è il solo. Molte altre aree sono attive quando osserviamo un’azione (vedi Figura 1). Il ruolo delle altre aree e in particolare della corteccia temporale viene spesso sottostimato dai sostenitori della teoria motoria e relegato ad una funzione di mera analisi visuo-motoria di basso livello.
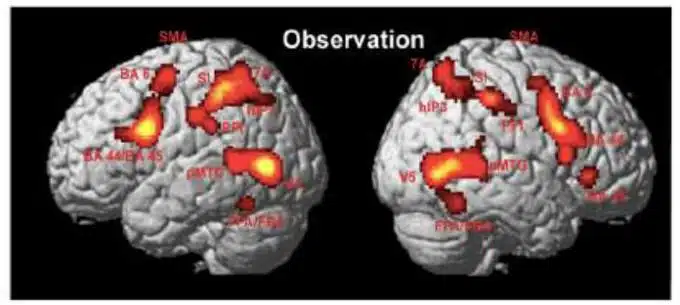
Aree che fanno parte dell’action-observation network (AON). Casper et al., 2010
Neuroni specchio e teoria cognitiva del riconoscimento delle azioni
I sostenitori della cosiddetta teoria cognitiva offrono una visione alternativa riguardo a come il nostro cervello associa i significati alle azioni. Punto chiave di questa ipotesi è che le rappresentazioni concettuali sono immagazzinate in aree prettamente concettuali che si trovano al di fuori del sistema sensorimotorio (Mahon e Caramazza, 2008; Papeo et al., 2009; Hickok, 2009). In altre parole, l’informazione semantica riguardante le azioni non dipenderebbe da un programma motorio specifico ma è astratta e si trova in regioni non-motorie.
Come abbiamo visto nel precedente paragrafo, le regioni motorie rispondono durante compiti che coinvolgono l’elaborazione di azioni. Rizzolatti e collaboratori (2001 p.6610) affermano che una
azione è compresa quando la sua osservazione causa una risonanza nel sistema motorio dell’osservatore.
I sostenitori della teoria cognitiva non negano che ci sia un’attivazione delle regioni motorie, ma rispondono che questa risonanza potrebbe essere la conseguenza di una connessione associativa con aree concettuali, o avere comunque altre funzioni meno specifiche. Questo significherebbe che quando osserviamo un’azione, prima viene attivata l’area concettuale non-motoria che contiene tutte le informazioni relative al significato dell’azione e che ci permette di comprendere l’azione, e solo in un secono momento, tramite connessioni associative, viene attivata l’area premotoria.
Studi neuropsicologici hanno esaminato le conseguenze di lesioni all’emisfero sinistro durante lo svolgimento di compiti in cui veniva chiesto di elaborare il significato di azioni. Se il riconoscimento e l’esecuzione di un’azione si basano sullo stesso meccanismo neurale, come sostiene la teoria motoria, allora entrambe le abilità dovrebbero essere compromesse quando le strutture neurali motorie e premotorie sono danneggiate (Pazzaglia et al., 2008). Al contrario, numerosi studi mostrano doppie dissociazioni tra riconoscimento di azioni e esecuzione di azioni (Rumiati et al., 2001; Negri et al., 2007; Kalénine et al., 2010; Urgesi et al., 2014). Questo significa che ci sono pazienti che non sono in grado di eseguire correttamente un’azione, ma sono però in grado di comprendere e interpretare le azioni altrui (Vannuscorps e Caramazza, 2016) e viceversa. Questi risultati vanno in direzione opposta rispetto a coloro che sostengono la teoria pura dei neuroni specchio. Questi studi, esaminando la connessione tra luogo della lesione e performance hanno individuato nella corteccia temporale e in particolare nel giro temporale mediale posteriore (pMTG) l’area concettuale dove sarebbe immagazzinata l’informazione concettuale delle azioni.
Questo risultato viene confermato da studi di fMRI che non solo mostrano l’attivazione di pMTG durante l’elaborazione di azioni, ma, tramite tecniche più avanzate, come il multivoxel pattern analysis (MVPA) mostrano come la corteccia occipito-temporale contenga informazione più astratta (e quindi la rappresentazione dell’azione distaccata dalla sua componente motoria), mentre il giro precentrale informazione di più basso livello (come può essere la chinematica di un movimento, la sua direzione ecc… – Wurm e Oosterhof, 2013; Lingnau, 2015; Wurm et al., 2015). Inoltre, in un recente studio di TMS, è stato dimostrato che la perturbazione di pMTG (collocato nella corteccia occipitotemporale) porta ad una interruzione del processo di riconoscimento semantico dei verbi (Papeo et al. 2014) e un’interruzione delle connessioni tra pMTG e l’area premotoria.
Neuroni specchio: spazio alle teorie moderate
Gli studi sopracitati suggeriscono quindi che l’informazione concettuale delle azioni è astratta ed è rappresentata nel lobo temporale e non nelle aree motorie e premotorie come sostiene le teoria motoria.
Si è visto come la teoria motoria e la teoria cognitiva del riconoscimento delle azioni si basino su due diverse assunzioni: la teoria motoria sostiene che il contenuto concettuale è costituito dall’informazione che è rappresentata nel sistema sensorimotorio e che quindi l’attività dei neuroni specchio è fondamentale per riconoscere il significato di un’azione. La teoria cognitiva, invece, sostiene che le rappresentazioni concettuali sono simboliche e astratte e che sono immagazzinate in aree concettuali al di fuori del sistema sensorimotorio e più in particolare, nella corteccia occipitotemporale e che quindi l’attività dei neuroni specchio non è necessaria per riconoscere il significato di un’azione. Queste due teorie si collocano a due estremi opposti e tendenzialmente una esclude l’altra. Tuttavia, tra questi due estremi si trovano altre teorie più moderate che, sbilanciandosi più verso un estremo o verso l’altro, cercano di conciliare queste due visioni.